Living entities have a tightly regulated dynamic system, where majority of the energy expenditure goes for maintaining the homeostasis. Most of the biological processes such as growth, development, metabolism etc are regulated at the crucial control point of protein translation and degradation (rate of protein turnover). Studies on different organisms/model systems have suggested that transcriptome modifies itself dynamically in response to different cellular conditions. The translational regulation is a slightly independent add-on layer of control after that, hence, during the cell processes a linear relationship of transcriptome and translatome is relatively difficult to establish. The efficacious identification of lesser abundant proteins is of lower magnitude, albeit of having sub-femtomolar level of sensitivity of modern state-of-the- art Mass spectroscopic instruments because of a narrow dynamicity and sequencing speed.
Consequently, the pre-eminent idea of enhancing the sequencing reliability and broadening the dynamic range of protein identification is in conjunction with the development of strategies to minimize proteomic complexity by using protein fractionation and affinity-purifications before proceeding to MS analysis. This approach has been successfully applied to protein distributed in specific subcellular compartments or with low copy number. A quantitative determination of a specific set of proteins that are differentially expressed in different subcellular locations at a specific temporal scale can provide a plethora of information about a cell`s phenotype and biology. Hence, a cornucopia of approaches have been used and developed, such as, differential 2D gel electrophoresis, relative or absolute quantification using isotope-coded affinity tags or isobaric tags, SILAC (stable isotope labeling by amino acids in cell culture), quantitative proteome analysis using 14N or 15N media etc, to understand the kinetics of translatome in variable conditions.
Importantly, these approaches have been successful in developing insights for biomarker discovery, mechanisms of neurodegeneration, protein-protein interactions and dynamics. In the course of recent developments in translatome, post-translational modifications have emerged as an important modulator or a control point to regulate many biological processes. Hence it becomes imperative to explore these specific "tagging" of cellular proteins in the sequence of exploration of proteomics. Though, these tagged proteins have a very small sample size, which makes the study of selective enrichment of newly synthesized proteins relatively troublesome. Regardless, identification of newly synthesized proteins and their differential enrichment would strengthen a wide range of differential proteomic profiling tools which subsequently will enhance our understanding of spatio-temporal dynamics of translatome.
Along the same line, a novel approach has been developed called BONCAT (Bioorthogonal non-canonical amino-acid tagging) to particularly recognize a subpopulation of recently translated proteins, initially started by Nobel laureate and synthetic chemist K. Barry Sharpless in 2001 as "Click chemistry". The cardinal principle of BONCAT method is the higher sensitivity of small bioorthogonal chemoselective groups. These groups add an additional and distinct layer of chemical functionality to their target biomolecule. These chemically modified targets can further be attached with an exogenously delivered set of probes which alleviate their isolation or detection in an extremely selective manner.
The most common derivatives used for BONCAT are azides and alkynes for labelling of proteins, glycans, and lipids used in the process of translation or protein modification in the cells. BONCAT is broadly divided into two steps:
Step 1: Labeling of newly synthesized proteins: In this step, the recently translated proteins are tagged with azide-bearing artificial methionine amino acid (AHA, azidohomoalanine; or HPG, homopropargylglycine) providing a novel azide functional group, which confer the differentiating layer from the pool of pre-existing proteins containing normal methionine. It is important to know here that, in animal systems azides or alkynes are not biologically synthesized except in few unialgal cultures. Moreover, these derivatives are non-toxic and stable in the biological systems. AHA can behave as an efficient proxy for natural methionine (an essential amino acid) and can easily be accepted by methionyl-tRNA synthetase as a substrate and incorporated in the translating peptide sequence.
The important characteristic property of these functional groups is the bioorthogonality, which does not allow the cross reactivity of these derivatives with the biological components. The chemistry involves reaction of Azides with alkynes using copper-catalyzed azide-alkynecycloaddition (CuAAC) specifically (Click Chemistry), hence enable specific tagging of a selective pool of proteins for a particular timeline or experimental condition, containing AHA and HPG within the group of complex biomolecules (unlabelled).
NOTE: The substitution of methionine in experimental conditions by non-canonical amino-acids (ncAA) such as AHA and HPG, can be ensured and enhanced by incorporating a methionine-depletion step right before the AHA or HPG addition. Absence of methionine from the medium during the metabolic labelling reaction has been reported to improve the fraction of incorporation.

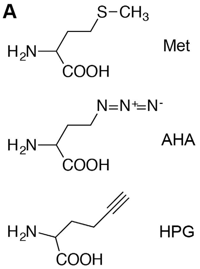
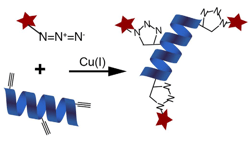
Figure 1: Azide and Alkyne derivatives used for BONCAT labelling.
Step 2: Tagging of AHA- and HPG-containing proteins: The AHA- and HPG-bound proteins can be covalently tagged with different reporters for different experimental designs such as fluorophores for imaging or biotin-Flag affinity tagging for enrichment studies. Addition of different derivatives of AHA and HPG allows pulse-chase experiments within a same experimental set up. Tagging of AHA and HPG labelled proteins with reporters enable studies of enriched newly synthesized proteins. Tagging of these proteins with bioorthogonal derivatives provide lesser complex samples promoting the identification of even those proteins that are expressed in low copy number.
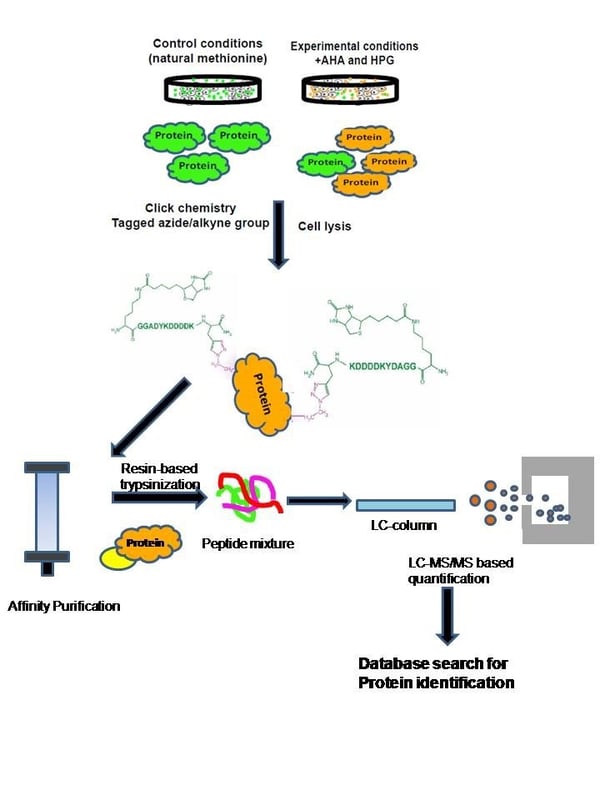
Figure 2: Workstation for BONCAT
BONCAT in different model systems
The critical part of developmental biology experiments is to maintain a very narrow and defined time window and be able to specifically label altered or enriched crucial proteins in that small developmental phase. One of the way used is analysing protein turnover rate using SILAC by incorporating labelled isotopes, categorically utilized in cell culture systems majorly. Although adult and embryonic murine tissues can be used too, it is a lengthy and convoluted process.
The most commonly and hassle free method for new protein labeling is AHA and HPG tagging, successfully used in-vitro (variety of cell lines, primary neuronal cells, organotypic slice cultures) as well as in zebrafish and Xenopus in vivo. It is also possible to perform global (feeding or Intraperitoneal injections) as well as local incorporation (intraocular injections) of AHA and HPG. The non-canonical amino acids (ncAAs) can easily integrate into different tissues of the juveniles or embryos if injected into the pregnant dams without compromising their developmental health. These methods present ncAA labelling (BONCAT) as a cutting edge tool to develop insights for the dynamics of protein synthesis and turnover in a temporal window of development or behaviour. Overall, AHA and HPG are not toxic and stable, therefore does not alter the rates of protein synthesis or degradation.
Advantages of BONCAT:
- BONCAT is not specifically limited to identification of recently synthesized proteins, but it also put forward a combination of different proteomic approaches in order to compare variability in different proteomes in just one MS experiment or between specific subpopulations of the translatome.
- Subcellular fractionation immunopurification of the modified complexes followed by BONCAT allows assessment of spatio-temporal dynamics of protein–protein interaction networks or protein enrichment in certain subcellular compartments, organelles.
- AGA and HPG tagged cell population with different reporters (fluorophores) can help gaining information about protein localization. Qualitative (fluorescence visualization) and quantitative (isolation and LC/MS/MS) analysis of protein dynamics present spatio-temporally defined snap shots or live imaging of the cellular systems.
- BONCAT allows identification of low abundance, low copy number newly synthesized proteins with higher magnitude as compared to SILAC.